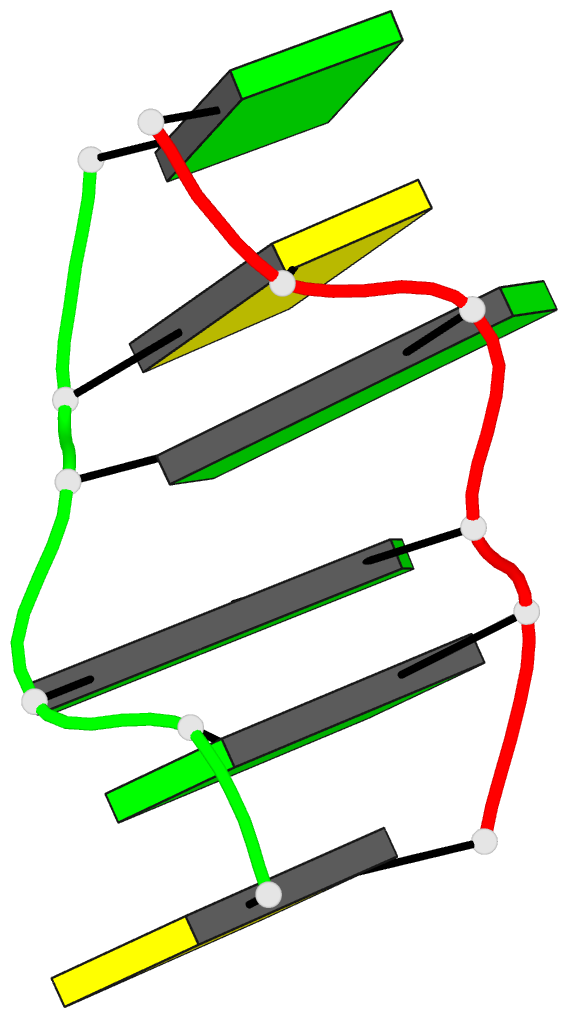
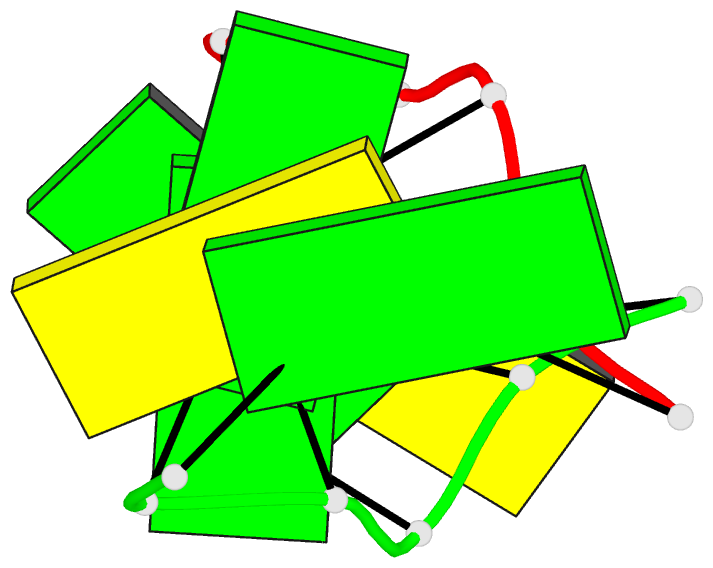
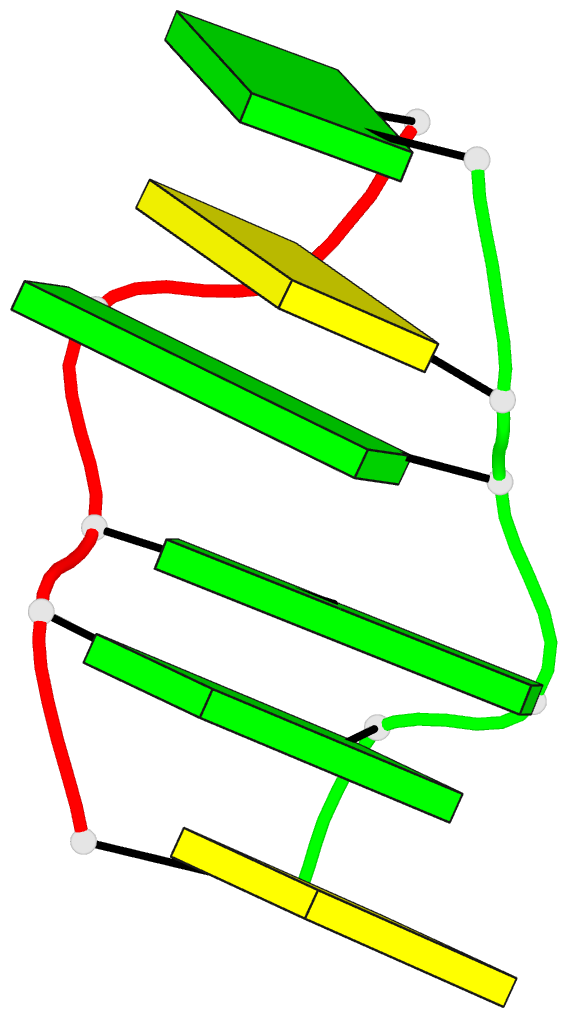
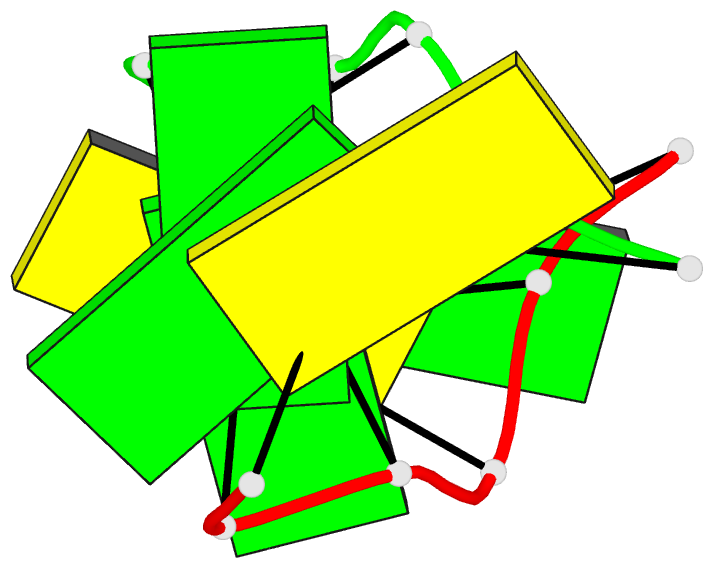
Summary information and primary citation
- PDB-id
-
145d;
SNAP-derived features in text and
JSON formats
- Class
- DNA
- Method
- X-ray (1.25 Å)
- Summary
- Structure and thermodynamics of nonalternating c-g base
pairs in z-DNA: the 1.3 angstroms crystal structure of the
asymmetric hexanucleotide d(m(5)cgggm(5)
cg)-d(m(5)cgccm(5)cg)
- Reference
-
Schroth GP, Kagawa TF, Ho PS (1993): "Structure
and thermodynamics of nonalternating C.G base pairs in
Z-DNA: the 1.3-A crystal structure of the asymmetric
hexanucleotide d(m5CGGGm5CG).d(m5CGCCm5CG)."
Biochemistry, 32, 13381-13392.
doi: 10.1021/bi00212a002.
- Abstract
- We have solved the single-crystal X-ray structure of
the complementary hexanucleotides d(m5-CGGGm5CG) and
d(m5CGCCm5CG). The hexamer duplex was crystallized as
Z-DNA, but contains a single C.G base pair that does not
follow the alternating pyrimidine/purine rule for Z-DNA
formation. This is the first crystal structure which serves
to illustrate the structural consequences of placing a
cytosine in the sterically disfavored syn conformation. In
addition, since these sequences are not self-complementary,
the individual strands of this asymmetric hexamer are
unique in sequence and therefore distinguishable in the
crystal lattice. Nevertheless, the crystal of this duplex
is isomorphous with other Z-DNA hexamer structures. The
asymmetry of this hexamer sequence required that the
structure be solved using two unique models, which are
distinguished by the orientation of hexanucleotides in the
crystal lattice. In one model (the GG model) the cytosine
in the syn conformation is packed against the terminal
guanine base of a symmetry-related hexamer, while in the
alternative model (the CC model) this cytosine sits exposed
in a solvent channel of the lattice. We find that neither
model alone can completely account for the observed
electron densities. The two models ultimately were refined
together. A composite structure consisting of 65% GG model
and 35% CC model refined to an R-factor of 19.3%, which was
significantly lower than refinements using either model
alone. A detailed analysis of these two structures shows
that, in spite of the out-of-alternation C.G base pair, the
features characteristic of Z-DNA have been maintained. Both
models, however, show significant local structural
adjustments to accommodate the single cytosine base which
is forced to adopt the syn conformation in each hexamer. In
general, it appears that in order to relieve the
energetically unfavorable steric contacts between the
cytosine base in the syn conformation and the deoxyribose
sugar, the base is forced into a highly buckled
conformation, and that this large buckle in turn alters the
conformation of neighboring residues. This unusual
conformation also significantly weakens base-stacking
interactions between the cytosine in syn and the adjacent
residues in the helix and affects the exposure of the bases
to solvent. We conclude that this crystal structure
provides a molecular rationale for why nonalternating bases
are energetically disfavored in Z-DNA.